Hormones at the tissue level
Hormones are essential to healthy functioning.1 Yet measuring hormone levels accurately at the tissue level continues to present a challenge. Serum levels of a hormone may rise faster than tissue levels (for instance, insulin in skeletal muscle1,2).
Many factors affect the passive and active delivery of hormones to tissues. For instance, localized inflammation can lead to increased transcellular gaps that increase permeability and allow hormones into cells more rapidly.3 Sex steroids, particularly estradiol, also alter vasodilation and endothelial function.4 Many sex hormones affect nitric oxide synthesis.5
Endothelium: An endocrine organ?
The endothelium is not uniform across all blood vessels in the body – it can be intermittent, continuous, thick, or thin.2 Importantly, the endothelium is not merely inert cells. Rather, the endothelium has dynamic, ongoing effects that are constantly responding to the environment.1,6
Some authors even call the endothelium “an endocrine organ.”7 They explain:
“The endothelium actively controls the degree of vascular relaxation/constriction, the extravasation of solutes, fluid, hormones, and macromolecules.”7 (emphasis added)
The endothelium acts as a barrier between blood and tissue, modulating the delivery of molecules like hormones. Even as a selectively permeable barrier, the endothelium is not uniform. Rather, this barrier permeability fluctuates over time and throughout the body.8
This fluctuation is important because the endothelium is “an important rate-limiting determinant of hormone and substrate access to target tissues.”2 This barrier function underlies the endothelium’s ability to regulate hormone access to the target tissues.2
Yet the argument for the endothelium as an endocrine organ is more complex. The endothelium is well known to harbor and release a variety of vasoactive peptide hormones, growth factors, coagulation factors, and more.9 It plays an essential role in vascular homeostasis, particularly with the vasodilators prostacyclin and nitric oxide and the more recently discovered endothelium-derived hyperpolarizing factor.10
In this article, we focus in on the microvasculature and tissue perfusion for hormones.
Microvasculature as the delivery site
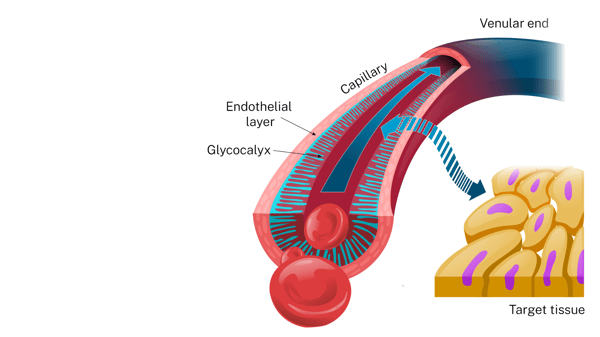
The capillaries of the microvasculature are the primary site where hormones cross the endothelium into the target tissues.11,12
The endothelium contains hormone receptors, such as estradiol13 and progesterone receptors,14 both of which can influence endothelial permeability.
The microvasculature is a dynamic network, constantly being remodeled in response to blood sugar, shear stress, inflammation, and more.15 Not all capillaries are used all the time. Rather, capillaries are recruited and remodeled as needed, based on a wide range of factors.16 Hormones are one of the factors that alter capillary recruitment, as well as anything that affects vasodilation.1 For example, insulin alters capillary recruitment.17Human studies have also confirmed that estrogen and progesterone also affect capillary perfusion.18
Inadequate capillary recruitment and resulting low tissue perfusion is correlated with and occurs early in the development of a number of conditions, including obesity, diabetes, dyslipidemia, and hypertension.19 Inadequate capillary recruitment by necessity decreases tissue perfusion for any molecules, including hormones.
The Endothelial Glycocalyx: An essential barrier
Within the endothelium, the micro-thin, gel-like endothelial glycocalyx (EGX) is the “primary molecular sieve” governing permeability.20 Every artery, vein, and capillary is lined with the protective shield of the EGX. The EGX is made up of glycoproteins and polysaccharides, and it is a semipermeable barrier between the blood and endothelial cells.
Much is still needed to learn about how the EGX functions in the microvasculature. This important area merits further research. A series of simulation studies have concluded that “our simulations suggest the presence of a semipermeable, space-filling glycocalyx layer in the vessel lumen is sufficient to promote homogeneity of microvascular perfusion.”21
When the EGX degrades, shedding pieces and becoming thinner, a range of negative effects occur:16
- Breakdown of sieve functionality (loss of oncotic pressure gradient)
- Widening of endothelial clefts
- Increased leukocyte and platelet adherence
- Reduction in microvascular outflow
This makes a strong case that EGX degradation impairs transport and delivery of molecules in the microvasculature. In animal models, this seems to hold true.
A few studies have directly looked at this question. Back in 1985, a hamster study showed that perturbation of the EGX led to reduced capillary perfusion.22 A rat study in 2011 demonstrated that as the EGX degrades, significant changes can be observed:23
- Decreased capillary volume
- Increased white cell adhesion
- Reduced capillary resistance.
A compromised EGX in the microvasculature over time very likely leads to compromised microvasculature. Disruption of the EGX also leads to endothelial hyperpermeability, which is correlated with a range of negative outcomes.24
Takeaways
Although more research is needed, it appears that protecting the EGX also protects the endothelium and enables proper delivery of hormones to target tissues. As a potential endocrine organ, protecting the endothelium may help patients with a range of hormone conditions.
Learn more about women's hormones and the vascular system in the next article.

References
1 Kolka CM, Bergman RN. The Barrier Within: Endothelial Transport of Hormones. Physiology (Bethesda). 2012;27(4):237-247. doi:10.1152/physiol.00012.2012
2 Yoon N, Dang TQ, Chasiotis H, Kelly SP, Sweeney G. Altered Transendothelial Transport of Hormones as a Contributor to Diabetes. Diabetes Metab J. 2014;38(2):92-99. doi:10.4093/dmj.2014.38.2.92
3 Pi X, Xie L, Patterson C. Emerging roles of vascular endothelium in metabolic homeostasis. Circ Res. 2018;123(4):477-494. doi:10.1161/CIRCRESAHA.118.313237
4 Chow RWY, Handelsman DJ, Ng MKC. Minireview: rapid actions of sex steroids in the endothelium. Endocrinology. 2010;151(6):2411-2422. doi:10.1210/en.2009-1456
5 Duckles SP, Miller VM. Hormonal modulation of endothelial NO production. Pflugers Arch. 2010;459(6):841-851. doi:10.1007/s00424-010-0797-1
6 Curry FR, Adamson RH. Tonic regulation of vascular permeability. Acta Physiol (Oxf). 2013;207(4):628-649. doi:10.1111/apha.12076
7 Krüger-Genge A, Blocki A, Franke RP, Jung F. Vascular Endothelial Cell Biology: An Update. Int J Mol Sci. 2019;20(18):4411. Published 2019 Sep 7. doi:10.3390/ijms20184411
8 Stevens T, Garcia JG, Shasby DM, Bhattacharya J, Malik AB. Mechanisms regulating endothelial cell barrier function. Am J Physiol Lung Cell Mol Physiol. 2000;279(3):L419-422. doi:10.1152/ajplung.2000.279.3.L419
9 Baumgartner-Parzer SM, Waldhäusl WK. The endothelium as a metabolic and endocrine organ: its relation with insulin resistance. Exp Clin Endocrinol Diabetes. 2001;109 Suppl 2:S166-179. doi:10.1055/s-2001-18579
10 Garland CJ, Hiley CR, Dora KA. EDHF: spreading the influence of the endothelium. Br J Pharmacol. 2011;164(3):839-852. doi:10.1111/j.1476-5381.2010.01148.x
11 Yuan SY, Rigor RR. Structure and Function of Exchange Microvessels. Morgan & Claypool Life Sciences; 2010. Chapter 2. Accessed November 5, 2020. https://www.ncbi.nlm.nih.gov/books/NBK54123/
12 Kolka CM. The vascular endothelium plays a role in insulin action. Clin Exp Pharmacol Physiol. 2020;47(1):168-175. doi:10.1111/1440-1681.13171
13 Russell KS, Haynes MP, Sinha D, Clerisme E, Bender JR. Human vascular endothelial cells contain membrane binding sites for estradiol, which mediate rapid intracellular signaling. PNAS. 2000;97(11):5930-5935. doi:10.1073/pnas.97.11.5930
14 Goddard LM, Murphy TJ, Org T, et al. Progesterone Receptor in the Vascular Endothelium Triggers Physiological Uterine Permeability Preimplantation. Cell. 2014;156(3):549-562. doi:10.1016/j.cell.2013.12.025
15 Yuan SY, Rigor RR. Signaling Mechanisms in the Regulation of Endothelial Permeability. Morgan & Claypool Life Sciences; 2010. Chapter 5. Accessed November 11, 2020. https://www.ncbi.nlm.nih.gov/books/NBK54119/
16 Jacob M, Chappell D, Becker BF. Regulation of blood flow and volume exchange across the microcirculation. Crit Care. 2016;20. doi:10.1186/s13054-016-1485-0
17 Rattigan S, Bussey CT, Ross RM, Richards SM. Obesity, Insulin Resistance, and Capillary Recruitment. Microcirculation. 2007;14(4-5):299-309. doi:10.1080/10739680701282796
18 Stachenfeld NS, Keefe DL, Palter SF. Estrogen and progesterone effects on transcapillary fluid dynamics. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 2001;281(4):R1319-R1329. doi:10.1152/ajpregu.2001.281.4.R1319
19 Levy Bernard I., Schiffrin Ernesto L., Mourad Jean-Jacques, et al. Impaired Tissue Perfusion. Circulation. 2008;118(9):968-976. doi:10.1161/CIRCULATIONAHA.107.763730
20 Hu X, Weinbaum S. A New View of Starling’s Hypothesis at the Microstructural Level. Microvascular Research. 1999;58(3):281-304. doi:10.1006/mvre.1999.2177
21 McClatchey PM, Schafer M, Hunter KS, Reusch JEB. The endothelial glycocalyx promotes homogenous blood flow distribution within the microvasculature. Am J Physiol Heart Circ Physiol. 2016;311(1):H168-H176. doi:10.1152/ajpheart.00132.2016
22 Cabrales P, Vázquez BYS, Tsai AG, Intaglietta M. Microvascular and capillary perfusion following glycocalyx degradation. J Appl Physiol (1985). 2007;102(6):2251-2259. doi:10.1152/japplphysiol.01155.2006
23 Lipowsky HH, Gao L, Lescanic A. Shedding of the endothelial glycocalyx in arterioles, capillaries, and venules and its effect on capillary hemodynamics during inflammation. Am J Physiol Heart Circ Physiol. 2011;301(6):H2235-H2245. doi:10.1152/ajpheart.00803.2011
24 Ushiyama A, Kataoka H, Iijima T. Glycocalyx and its involvement in clinical pathophysiologies. J Intensive Care. 2016;4(1):59. Published 2016 Sep 8. doi:10.1186/s40560-016-0182-z
IMPORTANT NOTICE:
The information on this webpage is for licensed healthcare practitioner education only, and is not to be disseminated to the general public.